“The IL6 axis plays a pivotal role in both acute and chronic inflammatory responses, operating through two distinct pathways: classical signalling via a membrane-bound IL6 receptor and trans-signalling mediated by a soluble IL6 receptor (IL6R), which enables IL6 activity in cells lacking the membrane receptor.”
Inflammation is a double-edged sword. It defends the body against infection and injury, yet when it becomes chronic, it can accelerate aging and fuel the very diseases that shorten human life. For decades, scientists have observed that people with higher levels of inflammatory markers like interleukin-6 (IL6) and C-reactive protein (CRP) tend to have shorter lifespans. But the critical question has always been: does inflammation cause mortality, or does it merely reflect underlying disease?
The team’s investigation demonstrates that the IL6 inflammatory pathway has a direct causal impact on human survival—but with a surprising twist: two components of the same pathway pull in opposite directions.
The Method: Mendelian Randomization
To establish causation, the researchers employed Mendelian randomization (MR), a technique that uses genetic variants as natural experiments. Because genes are randomly assigned at conception and fixed throughout life, they are not subject to the confounding factors—such as lifestyle, diet, or socioeconomic status—that plague traditional observational studies.
The team analyzed genetic data from approximately 750,000 individuals of European ancestry, focusing on four inflammatory biomarkers: interleukin-6 (IL6), its soluble receptor (IL6R), C-reactive protein (CRP), and growth differentiation factor-15 (GDF15). The primary outcome was all-cause mortality over a median follow-up of 11.7 years, with secondary outcomes including cardiovascular events and cancer.
Key Findings: Opposing Forces in the IL6 Pathway
The results revealed a remarkable biological duality. Genetically higher levels of the soluble IL6 receptor (IL6R) were associated with a reduced risk of all-cause mortality (odds ratio 0.95 per 1-standard deviation increase; p = 0.007). Higher IL6R levels also lowered the risk of atrial fibrillation, coronary artery disease, stroke, and lung cancer.
In stark contrast, genetically higher levels of IL6 itself were linked to an increased risk of mortality (odds ratio 1.05; p = 0.002). These findings suggest that IL6 and IL6R are biological opposites: IL6 drives harm, while IL6R protects.
The protective effects of IL6R were consistent across multiple sensitivity analyses, with no evidence of pleiotropy (where genetic variants influence outcomes through unintended pathways). A cis-Mendelian randomization analysis restricted to variants within the IL6R gene locus confirmed the protective association, reinforcing the causal relevance of this pathway.
CRP and GDF15: Biomarkers, Not Drivers
Notably, neither CRP nor GDF15 showed any significant causal effect on mortality or cardiovascular outcomes. Despite their well-established epidemiological associations with disease, these markers appear to be downstream indicators of inflammation rather than active drivers. As the authors note, this distinction is critical: CRP and GDF15 may be useful for predicting risk, but they are not themselves targets for intervention.
The Biological Mechanism: Classical vs. Trans-Signaling
The opposing effects of IL6 and IL6R are explained by the unique biology of the IL6 pathway. IL6 signals through two distinct routes. Classical signaling occurs when IL6 binds to membrane-bound IL6 receptors on certain cell types. Trans-signaling, however, occurs when IL6 binds to soluble IL6 receptors (sIL6R), allowing it to act on cells that lack membrane-bound receptors—including vascular and myocardial cells.
The genetic variants associated with higher sIL6R levels shift the balance away from trans-signaling, effectively dampening the inflammatory effects of IL6 in cardiovascular tissues. This reduces vascular inflammation, endothelial dysfunction, and thrombotic risk—mechanisms that directly contribute to atrial fibrillation, coronary artery disease, and stroke.
Clinical Implications: A Precision Target for Prevention
These findings have direct implications for drug development. IL6 receptor antagonists such as tocilizumab are already approved for inflammatory conditions like rheumatoid arthritis and giant cell arteritis, and have shown survival benefits in severe COVID-19. The genetic evidence presented here suggests that targeting IL6R could be an effective strategy for preventing cardiovascular disease and reducing mortality in high-risk populations.
Importantly, the neutral findings for CRP and GDF15 argue against broad anti-inflammatory approaches that target downstream markers. Instead, precision targeting of the IL6 signaling pathway—specifically through modulation of trans-signaling—appears to offer a more focused and potentially safer therapeutic avenue.
Limitations and Future Directions
The authors acknowledge several limitations. The analysis was restricted to individuals of European ancestry, which may limit generalizability to other populations. Additionally, while the study identified cardiovascular mechanisms as key mediators of IL6R’s mortality benefits, other potential pathways—such as metabolic or inflammatory diseases—remain to be explored.
Future research should focus on validating these findings in more diverse populations and conducting dedicated cardiovascular prevention trials with IL6R antagonists. The long-term safety of such interventions also warrants careful evaluation.
Future Perspectives and Conclusion
This study does not merely confirm that inflammation matters for longevity. It goes further, identifying a specific molecular axis—IL6 and its receptor—as a causal driver of human survival, with one component harming and the other protecting.
The perspective that emerges is one where the immune system’s inflammatory machinery can be precisely tuned. Rather than broadly suppressing inflammation—which could impair host defense—targeting IL6 trans-signaling offers a way to reduce cardiovascular risk while preserving essential immune functions.
As the authors conclude, “These results support IL6R antagonism as a potential strategy for cardiovascular disease prevention.” In an era where cardiovascular disease remains the leading cause of death globally, this genetic evidence provides a clear roadmap for translating inflammation biology into clinical practice.
Click here to read the full research paper published in Aging-US.
___
Aging-US is indexed by PubMed/Medline (abbreviated as “Aging (Albany NY)”), PubMed Central, Web of Science: Science Citation Index Expanded (abbreviated as “Aging‐US” and listed in the Cell Biology and Geriatrics & Gerontology categories), Scopus (abbreviated as “Aging” and listed in the Cell Biology and Aging categories), Biological Abstracts, BIOSIS Previews, EMBASE, META (Chan Zuckerberg Initiative) (2018-2022), and Dimensions (Digital Science).
Click here to subscribe to Aging-US publication updates.
“The idea of slowing, or even reversing, human aging has long occupied both science and imagination.”
Interest in healthier, longer lives is rising, supported by recent scientific advances in aging research. But turning those discoveries into everyday healthcare solutions remains a work in progress. In this landscape, longevity clinics have attracted attention as personalized alternatives to traditional medicine.
What Are Longevity Clinics?
Longevity clinics are private centers offering tailored programs designed to improve long-term health and slow biological aging. Using advanced diagnostics such as genetic sequencing, full-body imaging, and blood tests, they develop personalized plans that may include exercise, nutrition, hormone therapy, or experimental treatments. Frequently found in countries like the United States, Switzerland, and the United Arab Emirates, these clinics reflect a growing global interest in preventive healthcare, though their high costs and scientific credibility remain subjects of debate.
In this work, Dr. Demaria critically examines the rapid rise of longevity clinics, offering a thoughtful perspective on their current role, their potential to influence the future of healthcare, and the barriers they face in becoming credible contributors to aging science. He outlines both the opportunities these clinics present and the concerns surrounding their practices and impact.
Challenges
Longevity clinics aim to shift healthcare from treating illness to preventing it. Their appeal is based on the promise of early detection and personalized care tailored to each individual. However, these services often come at a significant cost, with some programs ranging from €10,000 to over €100,000 per year. This makes them accessible primarily to a small, wealthy segment of the population. As a result, concerns about fairness arise, especially considering that those most vulnerable to age-related health decline are often the least able to afford such care.
Opportunities
Despite the challenges, the editorial points out important contributions that longevity clinics could make. By collecting long-term data from clients, they may help researchers identify early warning signs of aging and detect age-related diseases earlier. Unlike traditional clinical trials, which are often short and disease-focused, these clinics track a broad range of health measures over time. When paired with artificial intelligence tools, this data could reveal meaningful patterns and support the development of better aging interventions.
The healthcare model promoted by longevity clinics also encourages people to actively manage their health, promoting lifestyle changes known to support healthy aging. Clinics often adopt new technologies and diagnostics faster than traditional institutions, potentially accelerating the translation of research into real-world use.
Concerns
Still, serious limitations remain. Some clinics offer therapies that are not well tested or not yet proven to be safe. Others provide test results that are difficult to interpret, and the lack of standardized protocols across clinics makes it harder to ensure consistency or accuracy. Tools like biological age calculators or hormone therapies may lack clear clinical value, which can lead to advice that is confusing or unsupported by strong evidence. Additionally, commercial motivations can outweigh scientific rigor. Furthermore, many clinics operate outside traditional healthcare systems, avoiding regulatory oversight. This not only creates safety concerns but also poses a risk to the credibility of the broader field of aging science.
Potential and Path Forward
What sets longevity clinics apart is their focus on personalization, prevention, and ongoing care. With greater scientific integration and ethical standards, they could become important partners in transforming how we approach aging and chronic disease. But for this to happen, certain conditions must be met.
The editorial outlines four key steps for the future. First, clinics should collaborate more closely with academic researchers and medical institutions. Second, testing protocols, biomarkers, and reporting methods must be standardized to improve consistency and scientific value. Third, broader access should be encouraged, whether through public health initiatives or insurance models. And fourth, there is a need to clarify the boundary between wellness services and medical care.
Conclusion
In summary, longevity clinics offer an idea of what future healthcare could look like: more personalized, preventive, and proactive. But without stronger scientific foundations, wider accessibility, and clear regulation, their promises may remain limited to a privileged few, leaving their full value uncertain. Whether they fulfill their promise will depend on continued collaboration with science. Equally important is a commitment to equitable, evidence-based care.
Click here to read the full editorial published in Aging-US.
___
Aging-US is indexed by PubMed/Medline (abbreviated as “Aging (Albany NY)”), PubMed Central, Web of Science: Science Citation Index Expanded (abbreviated as “Aging‐US” and listed in the Cell Biology and Geriatrics & Gerontology categories), Scopus (abbreviated as “Aging” and listed in the Cell Biology and Aging categories), Biological Abstracts, BIOSIS Previews, EMBASE, META (Chan Zuckerberg Initiative) (2018-2022), and Dimensions (Digital Science).
Click here to subscribe to Aging-US publication updates.
“This collection is published in memory of Professor Judith Campisi, a pioneering force in the field of cellular senescence whose groundbreaking work shaped the understanding of senescence in aging, cancer, and tissue homeostasis.”
BUFFALO, NY — May 1, 2025 —Aging (Aging-US) invites submissions for a Special Collection dedicated to the theme of cellular senescence, spanning its basic mechanisms, physiological and pathological functions, and clinical applications.
This collection is published in memory of Professor Judith Campisi, a pioneering force in the field of cellular senescence whose groundbreaking work shaped the understanding of senescence in aging, cancer, and tissue homeostasis. Her legacy continues to inspire generations of scientists working to decode the complex biology of senescent cells and their impact on health and disease.
We welcome original research articles, reviews, and perspectives on topics including:
Fundamental mechanisms of senescence induction and maintenance
Regulation and context-specific roles of the senescence-associated secretory phenotype (SASP)
Beneficial and detrimental effects of senescent cells in vivo
Senescence in development, aging, regeneration, and age-related diseases
Biomarkers, imaging, and tools for senescence detection and quantification
Therapeutic targeting of senescent cells: senolytics, senomorphics, and clinical translation
This Special Collection is guest edited by Han Li and Irina Conboy, both internationally recognized leaders in the study of senescence and aging.
In a recently published Aging-US paper, researchers investigated potential biomarkers of severe COVID-19 among recovered patients.
The Trending With Impact series highlights Aging (Aging-US) publications that attract higher visibility among readers around the world online, in the news, and on social media—beyond normal readership levels. Look for future science news about the latest trending publications here, and at Aging-US.com.
—
Listen to an audio version of this article
Among people who have been fortunate to recover from COVID-19, at least 40% had or have long-term lingering effects from this disease. Frequently appearing months after recovery, these “long covid” effects can include (not limited to) fatigue, trouble sleeping, difficulty concentrating, joint or muscle pain, and respiratory issues, such as shortness of breath and chest pain. Researchers are still unsure as to whether or not lasting COVID-19 effects in the lungs are associated with the severity of disease at the time of infection.
“Thus, prospective studies related to outcomes following recovery from COVID-19 might improve our understanding of this disease, its sequelae, and possible interventions to improve this situation.”
A total of 108 recovered COVID-19 patients admitted to hospitals throughout Spain (46.2% of whom had severe cases) were enrolled in this study. Lung function was measured by the capacity to diffuse carbon monoxide. Samples of serum and induced sputum (phlegm) were collected from the patients and used to evaluate the relationships between patients with residual inflammation in the lungs. Within these samples, the researchers analyzed and compared the levels of 92 protein biomarkers, including various chemokines, cytokines, growth factors, interleukins, and the CUB domain-containing protein 1 (CDCP1)—a cell surface glycoprotein. High levels of CDCP1 were previously observed in some severe cases of COVID-19 in children.
“In COVID-19-infected children who developed acute vasculitis, CDCP1 was one of the most significantly upregulated genes [25], but this complication was not observed in our study.”
The Results
Of all 92 biomarkers, multivariate analysis showed only elevated levels of serum CDCP1 in individuals recovering from severe COVID-19. To their surprise, they also found a positive relationship between CDCP1 and TGFb1 in sputum samples (irrespective of severity). The researchers found a significant difference in lung function (as measured by diffusing capacity for carbon monoxide (DLCO)) between those who had severe cases of COVID-19 and those who had mild/moderate cases. Differences in serum proinflammatory cytokines were not observed between the two groups of recovered COVID-19 patients, indicating that these biomarkers subside after recovery. Correlations between serum and induced sputum levels were detected for only a few biomarkers.
“Independent predictors of severe disease were DLCO <80% and the serum CDCP1 value.”
Conclusion
The authors were forthcoming about the limitations of their study. The team did not use a control group, although, the goal of this study was to investigate the severity of COVID-19. The cohort was relatively small, and the researchers note the need to further study the role of CDCP1. They also suggest that prospective studies should follow patients in recovery from COVID-19 in order to continue improving our understanding of this still novel virus. Such information could aid in the development of interventions to improve patient prognoses long-term.
“In conclusion, although the long-term impact of high serum levels of CDCP1 is still unknown, we should be alert to the potential implications for lung disease. For this reason, it is necessary to follow such patients for longer periods of time to detect and adequately treat potential pulmonary sequelae.”
Click here to read the full research paper published by Aging (Aging-US).
Aging (Aging-US) is an open-access journal that publishes research papers bi-monthly in all fields of aging research. These papers are available to read at no cost to readers on Aging-us.com. Open-access journals offer information that has the potential to benefit our societies from the inside out and may be shared with friends, neighbors, colleagues, and other researchers, far and wide.
Behind the Study is a series of transcribed videos from researchers elaborating on their recent oncology-focused studies published by Aging. A new Behind the Study is released each Monday. Visit the AgingYouTube channel for more insights from outstanding authors.
Well, most of my lab works on a process called cellular senescence, which is a cellular response to stresses and damage, many of which increase with age. And it’s now clear from mouse models that if you eliminate senescent cells, which increase with age, you can increase the health span of a mouse – not necessarily the lifespan, but the health span. So it becomes kind of important to have ways of identifying senescent cells in detail, and we have not been able to do that so far with absolute certainty because there frankly are no senescent-specific markers. So there are markers that are commonly expressed by senescent cells, but none of them are absolutely specific.
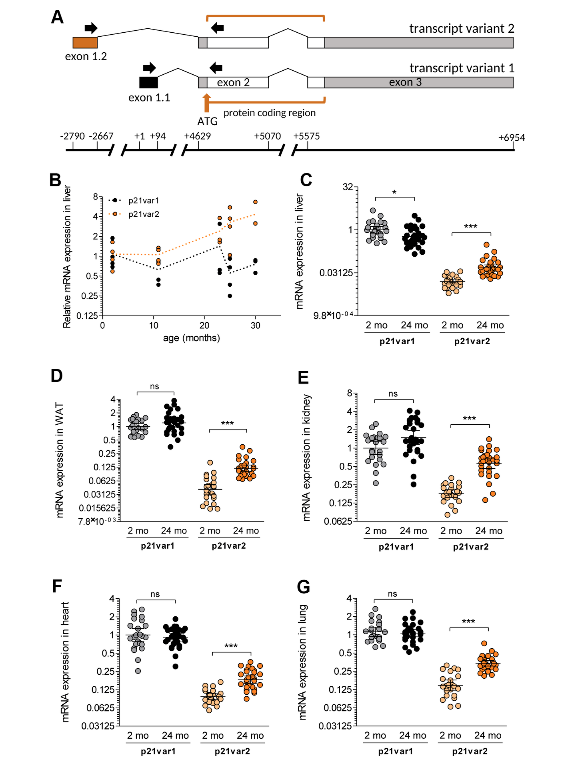
Figure 1. The Cdkn1a variant 2 transcript is preferentially induced during aging.
And so what we have done is we have looked at one of those markers, which is a gene called Cdkn1a and it codes for approaching, called P21. So everyone knows that P21 is one of those common biomarkers of aging, but it also is not necessarily strictly limited to aging. And what we’ve found is that there are two mRNAs that are made from that gene, that had been known before. We looked at these two mRNAs separately and found that one of them, which is called the variant 2, is a better marker of senescence and aging than the other mRNA. And that gives us a little bit of a edge in trying to unambiguously identify senescent cells in vivo and even in culture.
So the importance of this work is that it helps refine our ways of identifying these cells. We now know that these cells are important in aging, certainly in mice, probably in humans as well. So with this group of mine, many of which come from Spain or France or Russia, many of them contributed to refining this marker and allowing us to be able to have a better way of having some confidence that a senescent cell is indeed senescence.
And I can stop here.
Click here to read the full study published by Aging.
Aging is an open-access journal that publishes research papers monthly in all fields of aging research and other topics. These papers are available to read at no cost to readers on Aging-us.com. Open-access journals offer information that has the potential to benefit our societies from the inside out and may be shared with friends, neighbors, colleagues, and other researchers, far and wide.
Is there an association between biomarkers of aging and lung function? Researchers conducted a study which aimed to find out.
The Trending with Impact series highlights Aging publications that attract higher visibility among readers around the world online, in the news, and on social media—beyond normal readership levels. Look for future science news about the latest trending publications here, and at Aging-US.com.
—
Listen to an audio version of this article
By 2050, the United States Census Bureau estimates that 83.7 million people aged 65 years and older will be living in the U.S.—a number that will nearly double the 2012 estimated population. As the scale of the elderly population magnifies, additional aging research will continue to be increasingly relevant.
“According to the American Lung Association, the lung matures by age 20-25 years, and its function declines gradually after the age of 35 [30].”
Among the elderly population, lung function has been found to vary, even between those who have never smoked, are the same height, and of the same chronological age. This has led researchers to wonder if lung function decline is part of the underlying biological aging processes. No published studies had investigated the associations between epigenetic aging biomarkers and lung function, until 2020.
“In this present study, we hypothesized that some of these BoA are associated with lower lung function.”
The Study
From 1961 to 1970, healthy U.S. males between the ages of 21 and 81 enrolled in the ongoing Veterans Affairs Normative Aging Study. One of the objectives of the study is to characterize the biomedical and psychosocial parameters of normal aging (distinct from the development of disease). There are a total of 2,280 participants in the Normative Aging Study (NAS). In the current Aging study, researchers included 696 elderly men from the NAS.
“The present study included 696 elderly men with 1,070 visits during years of 1999-2013.”
In search of associations between biomarkers of aging and lung function, the researchers first collected the study participants’ personal characteristics, including age, smoking history, height, weight, BMI, education, blood work, and other measures. They then analyzed lung function using three tests: forced expiratory volume in one second (FEV1), forced expiratory volume in one second / forced vital capacity (FEV1/FVC), and maximum mid-expiratory flow (MMEF).
Next, the team analyzed the participants’ epigenetic biomarkers of age; including GrimAgeAccel, PhenoAgeAccel, intrinsic epigenetic age acceleration (IEAA), extrinsic epigenetic age acceleration (EEAA), and Zhang’s DNAmRiskScore; as well as non-epigenetic biomarkers of age, including telomere length and mitochondrial DNA copy number (mtDNA-CN). They then assessed for associations between these biomarkers and the three measures of lung function.
Conclusion
“In this longitudinal cohort of 696 elderly males, we found that GrimAgeAccel and Zhang’s DNAmRiskScore were associated with lower lung function, including FEV1, FEV1/FVC, and MMEF.”
The researchers found that the GrimAgeAccel and Zhang’s DNAmRiskScore were both associated with lower lung function in all three measures of lung function. They found no correlation between non-epigenetic aging biomarkers and lung function, but the researchers mention several limitations of their study. Their results suggest that epigenomic variation could help illuminate the pathogenesis of the reduced lung function that comes with age.
“Epigenetic mechanisms such as DNAm may provide further explanation for decreases in lung function as individual age.”
Click here to read the full paper, published by Aging.
Aging is an open-access journal that publishes research papers monthly in all fields of aging research and other topics. These papers are available to read at no cost to readers on Aging-us.com. Open-access journals offer information that has the potential to benefit our societies from the inside out and may be shared with friends, neighbors, colleagues, and other researchers, far and wide.
Researchers explain their studies that were published in Aging
Behind the Study is a series of transcribed videos from researchers elaborating on their recent oncology-focused studies published in Aging. A new Behind the Study is released each Monday. Visit the AgingYouTube channel for more insights from outstanding authors.
—
Greetings. My name is Andrei Gudkov. I am working in Roswell Park Cancer Institute, designated cancer center located in Buffalo, New York. I am Senior Vice President for Basic Science and chair of Department of Cell Stress Biology. My research is focused on understanding of the mechanisms of deregulation of a variety of stress response pathways in cancer cells as well as in normal cells in relation to cancer origin, progression, or engraftment and trying to use the information which we are generating during this research to come up with new types of treatment of cancer or cancer prevention.
Recently, our interests have significantly switched towards studying of the mechanisms of aging in its relations to cancer, since, as we all know, both conditions are closely connected. During the last, probably 20 years, one of the central theories of aging in mammals has been evolving towards connection between chronic sterile inflammation, which is accumulating in tissues with age of a mammal, including humans, with systemic decline in regeneration capabilities, in function of organs and tissues, and increasing risk of major diseases, altogether known as aging-related diseases. And the source of this inflammation, its origin, has been the central focus of studies of many.
During last couple of years, the dominating opinion in the field is about the central role of senescent cells, cells which chose to stay irreversibly growth-arrested in response to DNA damage, which they acquire during their life. And, through that, change their phenotype in more significant way than just growth arrest, acquiring the ability to secrete a spectrum of pro-inflammatory factors.
These senescent cells, which initially were defined as such in tissue culture experiments, eventually were proclaimed to be the main suspects in their putative role of inflammation creators in aging organism. This idea has become really popular, especially following a series of brilliant works coming from number of laboratories, in which senescent cells were detected in vivo in mice and mouse models. And when these mice were treated with agents which eradicated these senescent cells, numerous signs of rejuvenation were observed.
I’m talking about the first paper of that kind appeared in 2011, Mayo Clinic, and the group led by Jim Kirkland and Jan van Deursen and a series of follow-up papers with similar results. In general, the idea of putting senescent cells in the position of the key sources of sterile chronic inflammation associated with aging came from Judy Campisi, who has provided the most important discoveries in that field.
Well, this theory is extremely appealing for many reasons. First, it is very well supported by evidence. Indeed, senescent cells, when they turn into senescents in culture, switch their phenotype into, so-called, SASPs, and that’s an associated secretory phenotype, the state in which cells continuously secrete pro-inflammatory factors. Second, these cells appear in culture as a result of serial passaging resembling aging. And, therefore, this link became kind of natural between aging and senescent cells. The presumption was that certain cells in the body who used up the number of divisions they can go through before they reach this state may be increasing with age and, therefore, these cells accumulate.
Each of them may become the source of sterile inflammation. Each single one provides a very weak signal, but, when they accumulate altogether, the impact may become significant and translated into pathological conditions. So recently, there were very few – and, even now, it is like that – very few biomarkers of senescent cells, none of which is very reliable because every single biomarker is kind of promiscuous and is not universally selective for senescent cells.
Among these biomarkers, two have been most popular. One is high level of expression of, so-called, senescent-associated senescent-associated beta-galactosidase, which can be detected chemically in fixed cells and tissues which undergo staining, including X-gal, which turn beta-galactosidase reaction into the blue dye under conditions which is not optimal for endogenous beta-galactosidases mammalian cells at low pH. And, under these conditions, the background beta-gal activity of normal cells is practically not seen and senescent cells become brightly visible. So this reaction, which unfortunately requires a cell… It can not be done on paraffin-embedded sections and require preservation of the enzymatic activity and, therefore, is available, mostly, on the frozen sections or in cells in culture… has been used very, very frequently. And in many papers, it has been just the only assay which was used for detection of so-called senescent cells.
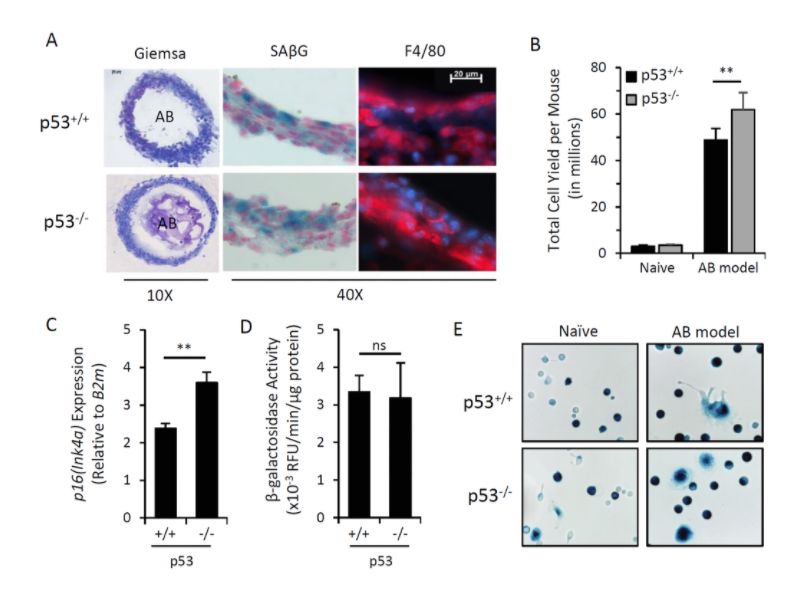
Figure 1.Induction of p16Ink4a and SAβG in macrophages does not require p53.
The other biomarker, which resulted from a detailed analysis of promoters which are active selectively in senescent cells is the gene encoding cyclin-dependent kinase inhibitor p16. And the genes name is INK4a. In fact, this promoter of this gene is frequently upregulated in senescent cells, and it has relatively low background in other cells of the organism.
Again, p16 activation is not limited to senescent cells and, moreover, not every senescent cells has elevated p16, but that’s the best we have as of today. That is why, whenever the investigators want to create a mouse model in which they could have the desirable gene expressed exclusively in senescent cells, they use p16 promoter. And there are several mouse models; I’m aware of three in which reported constructs were put under p16 promoter. And the claim was that, when these reporters become obviously expressed in mouse tissues, that was interpreted as accumulation of senescent cells. Also, one can put under this promoter the gene which enables selective eradication of cells with this expression, and, therefore, there is an opportunity to selectively kill such cells. Again, this can be interpreted as a selective eradication of senescent cells.
Using these models, two groups of investigators claim that eradication of senescent cells in aged mice led to substantial demonstration of signs of rejuvenation and, in one case, with increased lifespan. Well, obviously, these data not only provided a very powerful support for the theory about the role of senescent cells in aging but also provided the proof of concept for development of pharmacological approaches to anti-aging treatment and treatment of conditions which lead to the high risk of development of age-related diseases, including cancer.
We obtained such mice in our laboratory, and we have been working with them during last couple of years. The mice we are using are coming from the laboratory of Norman Sharpless from North Carolina. And they have a luciferase reporter gene, which is substituting one of the alleles of p16 and, thereby, being expressed from the p16 promoter. We were pleased to see that these mice accumulate p16-driven luciferase-positive cells detected by in vivo imaging during their lives, which, actually, very well fit the senescent cell theory in their accumulation during life.
However, we were very surprised not seeing accumulation of these cells following total-body radiation or treatment with other genotoxic conditions, which, supposedly, should create lots of senescent cells. We also were puzzled that we were unable to see activation of p16-driven luciferase when we take tissues from these mice and isolated mesenchymal cells from these tissues in vitro and then turn them into senescents, and we failed to see activation of luciferase.
Again, all this together stimulated us to look at the nature of p16-positive cells in these mice and determined their nature, their origin, and their fate in vivo. We started from following the consequences of injection of cells, which would turn into senescents in vitro following injection in vivo into mice. And we labeled cells. We made cell senescents in culture by gum radiation. Then, we injected them intraperitoneal, subcutaneously, into mice. And we looked for their presence by monitoring the label which they were marked with.
Well, it appears that these labeled cells – their traces are disappearing quite quickly, and, within a few days, there are none left in the mice. However, if you put normal cells of similar origin, they actually last much longer. That was the first indication that there may be a mechanism of selective eradication of senescent cells in the body. To check this mechanism and one of our hypotheses was that this mechanism is associated with physical attack of some cells of immunity against senescent cells, and there’s supposed to be innate immunity because it’s happening immediately without any education over the organism.
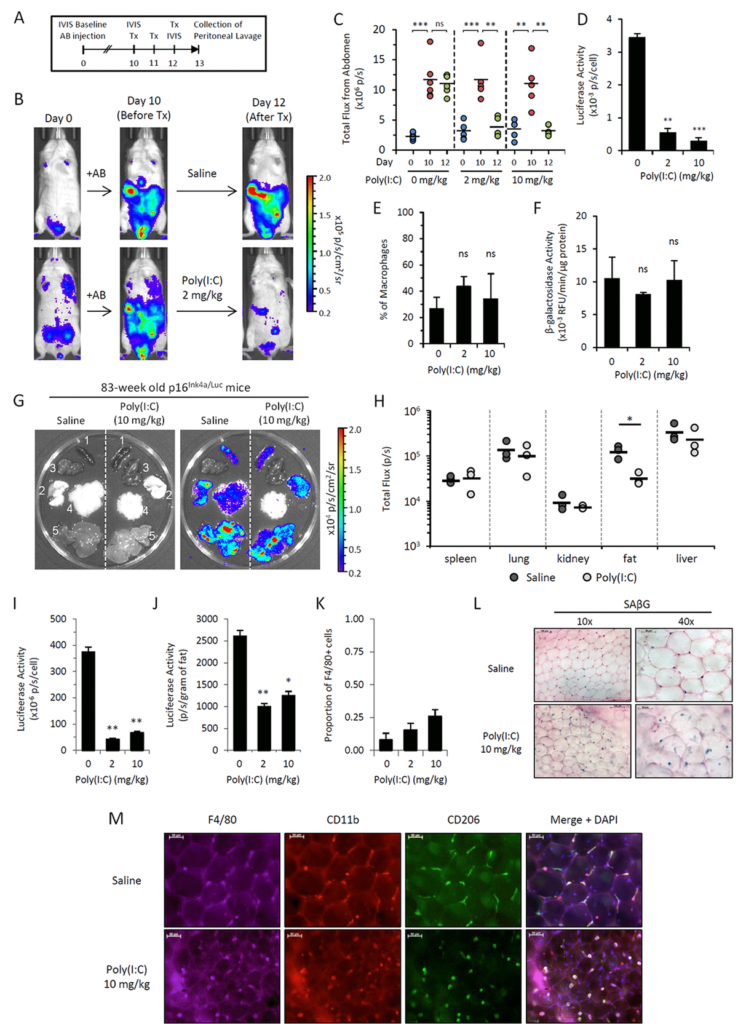
Figure 5.Poly(I:C) abrogates elevated p16Ink4a expression in two independent in vivo models.
We use a trick in which we embedded senescent cells created in vitro into algenate beads, small spheres consisting of a polymer, which enables to keep cells alive inside them, does not interfere with acquisition of nutrients and oxygen by the cells, but prevents any attack against the cells from any immunocytes. When we took these beads filled with senescent cells and put them in peritoneal cavity of mice, we were pleased to see that they are lasting four weeks without significant death, indicating that senescent cells, who disappear if they are injected without protective beads, are indeed killed by some, so far, unknown mechanism.
In order to identify the executors of senescent cells, we put these beads filled with senescent cells as bait inside, very peritoneal cavity of normal mice, and two weeks later, we pulled them out and analyzed who was accumulating in terms of how cells around these beads in lavage liquid, as well as in the capsule, which was formed around every bead.
Our results brought us to a very important and quite striking observational. The major part of the cells, which was so in these beads as well as in the lavage, appeared to be cells with macrophageal markers on them, which appeared to be bright fluorescence, meaning that they have activated p16, and also positive for beta-gal staining conducted under conditions we are using to reveal senescent cells. So we had to conclude that senescent cells put in the beads attract, probably, by the products of their secretion special subtypes of immune cells, significant proportion of which become reprogrammed to start expressing two biomarkers which people have been using to distinguish senescent cells.
We studied these macrophages in detail, and, after we published our first paper in which we describe this phenomenon, we published a second one, also in Aging, where their properties were described in further details. And we confident that these are bonafide macrophages, not only because they have have biomarkers, they have surface antigen specific for macrophages, but also they are capable of phagocytosis and, moreover, they can be selectively killed by liposome-embedded clodronate, a poison which only kills cells capable of phagocytosis. This killing could be done both in vitro and in vivo when you inject liposomal clodronate inside mice.
So, as far as the presence of these cells in the body of those mice which are not embedded with algenate beads with senescent cells, today, we are confident that these macrophages are accumulating in subcutaneous fat of aged mice in large numbers. And, again, they express biomarkers of macrophages that can be selectively eradicated by clodronate.
So, altogether, it means that the cells which become p16-positive vivo, not necessarily our senescent cells – our operations does not disprove that the signal which we and other investigators are seeing in these mice and increasing with age is not associated with senescent cells. So, potentially, certain proportion of cells we see are, indeed, senescent. However, we are confident that significant part of the signal goes from macrophages, which can be induced into the phenotype associated with expression of both senescent markers when they’re exposed to senescent cells. What is also interesting that this phenotype is reversible. And, in our second paper, we provide a number of physiological stimuli which can either stimulate or suppressed acquisition of this phenotype by macrophages.
All this, together, provides a very interesting step forward in evolution of the theory of aging associated with accumulation of certain specific cell types, contributing to the sterile inflammation occurring in tissues. Today, we can say that those cells which we claim to be the main source of that are not necessarily senescent, but also can be immunocytes who share with senescent cells some of their properties but are not senescent by nature and simply reprogrammed macrophages.
What is the relative impact of these macrophages versus senescent cells towards the process of aging is a very important question, not only from a theoretical standpoint, but also from practical standpoint because, from the time when senescent cells were claimed to be the key players of aging, there have been a substantial effort in the field in generating and testing senolytic compounds, drugs, emerging drugs, which potentially can have anti-aging effect due to eradication of senescent cells from the body.
Whether senolytic compounds would, indeed, solve the issue because, presumably, they will eliminate only a part of the p16-positive cells. To what extent, we need to redirect our attention to the senescent cell-associated macrophages as potential alternative source of secreted factors is an open question. And these are the questions which we are trying to address in our ongoing work, which stems from these observations. Thank you.
Click here to read the full study published in Aging.
—
Aging is an open-access journal that publishes research papers monthly in all fields of aging research and other topics. These papers are available to read at no cost to readers on Aging-us.com. Open-access journals offer information that has the potential to benefit our societies from the inside out and may be shared with friends, neighbors, colleagues, and other researchers, far and wide.